|
0.1.26
|
|
0.1.26
|
| Dependencies | None |
|---|---|
| Python Import | from pyhelios import PhotosynthesisModel |
| Main Class | PhotosynthesisModel |
| Dependencies | None |
|---|---|
| Platforms | Windows, Linux, macOS |
| GPU | Not required |
| Constructors |
|---|
| PhotosynthesisModel |
| Primitive Data Label | Symbol | Units | Data Type | Description | Available Plug-ins | Default Value |
|---|---|---|---|---|---|---|
| radiation_flux_PAR | \(Q\) | W/m2 | float | Radiative flux in PAR band. NOTE: this is automatically converted to units of photon flux density, which are the units used in the photosynthesis model. | Can be computed by RadiationModel plug-in. | 0 |
| temperature | \(T_s\) | Kelvin | float | Primitive surface temperature. | Can be computed by EnergyBalanceModel plug-in. | 300 K |
| air_CO2 | \(C_a\) | \(\mu\)mol CO2/mol air | float | CO2 concentration of air outside of primitive boundary-layer. | N/A | 390 \(\mu\)mol/mol |
| moisture_conductance | \(g_S\) | mol air/m2-s | float | Conductance to moisture between sub-stomatal cells and leaf surface (i.e., stomatal conductance). | Can be computed by StomatalConductanceModel plug-in. | 0.25 mol/m2-s |
| boundarylayer_conductance** | \(g_H\) | mol air/m2-s | float | Conductance to heat between leaf surface and outside of the boundary-layer (i.e., boundary-layer conductance). | Can be computed by BLConductanceModel plug-in, or by EnergyBalanceModel plug-in if optional output primitive data "boundarylayer_conductance_out" is enabled. | 1.0 mol/m2-s |
| twosided_flag | N/A | N/A | uint | Flag indicating the number of primitive faces with heat transfer (twosided_flag = 0 for one-sided heat transfer; twosided_flag = 1 for two-sided heat transfer). This value is retrieved from the primitive's assigned material when available (see Context::getMaterialTwosidedFlag()), with fallback to primitive data if no user-assigned material exists. | N/A | 1 |
| stomatal_sidedness | \(\zeta\) | unitless | float | Ratio of stomatal density on the upper leaf surface to the sum of the stomatal density on upper and lower leaf surfaces. Note: if "twosided_flag" is equal to 0, stomatal_sidedness will be automatically set to 0. | N/A | 0 |
**The photosynthesis model will also check for primitive data "boundarylayer_conductance_out" if "boundarylayer_conductance" does not exist. If you are using the energy balance model to calculate the boundary-layer conductance, you should enable optional output primitive data "boundarylayer_conductance_out" so that other plug-ins can use it.
| Primitive Data Label | Symbol | Units | Data Type | Description |
|---|---|---|---|---|
| net_photosynthesis | \(A\) | \(\mu\)mol CO2/m2-sec | float | Net rate of carbon transfer per unit one-sided area. |
Note: Optional output primitive data functionality is not yet implemented in PyHelios. In native Helios C++, this is done by calling PhotosynthesisModel::optionalOutputPrimitiveData(), which has no PyHelios equivalent at present.
| Primitive Data Label | Symbol | Units | Data Type | Description |
|---|---|---|---|---|
| Ci | \(C_i\) | \(\mu\)mol CO2/mol | float | Intercellular CO2 concentration. |
| Gamma_CO2 | \(\Gamma\) | \(\mu\)mol CO2/mol | float | Photosynthetic CO2 compensation point (including "dark respiration"). FvCB model only. |
| limitation_state | N/A | N/A | int | Photosynthesis limitation state. FvCB (C3): 0 = Rubisco-limited, 1 = electron-transport-limited. C4 (von Caemmerer 2021): 1 = enzyme-limited, 2 = electron-transport-limited. |
| Cm | \(C_m\) | \(\mu\)bar | float | Mesophyll cytosolic CO2 partial pressure. C4 model only (helios-core v1.3.72+). |
| Vp | \(V_p\) | \(\mu\)mol CO2/m2-sec | float | PEP carboxylation rate. C4 model only (helios-core v1.3.72+). |
PyHelios v0.1.21+ exposes the von Caemmerer (2021) steady-state C4 photosynthesis model (helios-core v1.3.72+). Activate it with setModelTypeC4(), then load species defaults from the C4 library or apply your own parameter array:
The 43-float coefficient array returned by getC4CoefficientsFromLibrary() / getC4ModelCoefficients() (and consumed by setC4ModelCoefficients()) is laid out as follows:
| Group | Slots | Fields |
|---|---|---|
Temperature-responsive rates (4 floats each: value_at_25C, dHa, Topt_C, dHd) | 0–19 | Vpmax, Vcmax, Jmax, Rd, gm |
| Rubisco + PEPC kinetic constants at 25 °C | 20–24 | Kc, Ko, Kp, γ*, Om |
| Activation energies (kJ/mol) | 25–29 | dH_Kc, dH_Ko, dH_Kp, dH_γ*, dH_Om |
| User-tunable scalars | 30–42 | α_psII_fraction, x_etr_partition, Vpr, Rm_frac, fcyc, gbs, ao, absorptance, f_spectral, θ_etr, h_protons, H_J, H_Jcyc |
Each temperature-responsive rate uses the same -1 sentinel convention as setFarquharMesophyllConductance — a negative dHa means "no temperature response", a negative Topt_C means "monotonic Arrhenius", a negative dHd means "default deactivation energy".
| Parameter | Symbol | Units | Default (Setaria) | Description |
|---|---|---|---|---|
| Temperature-responsive rates | ||||
| Vpmax | Vpmax | μmol/m²/s | 200 (dHa=50.1) | Maximum PEPC activity. Boyd et al. (2015). |
| Vcmax | Vcmax | μmol/m²/s | 40 (dHa=78.0) | Maximum Rubisco activity. Boyd et al. (2015). |
| Jmax | Jmax | μmol/m²/s | 247.69 (peaked, Topt=43, dHd=260) | Maximum linear ETR. Re-fit from von Caemmerer (2021) Gaussian. |
| Rd | Rd | μmol/m²/s | 1.0 (dHa=66.4) | Mitochondrial respiration. |
| gm | gm | mol/m²/s/bar | 1.0 (dHa=49.8) | Mesophyll conductance. Ubierna et al. (2017). |
| Kinetic constants (simple Arrhenius, edit struct fields directly) | ||||
| Kc_25, dH_Kc | Kc | μbar | 1210 / Ea=64.2 | Rubisco Michaelis constant for CO₂. |
| Ko_25, dH_Ko | Ko | μbar | 292 000 / Ea=10.5 | Rubisco Michaelis constant for O₂ (=292 mbar). |
| Kp_25, dH_Kp | Kp | μbar | 82 / Ea=38.3 | PEPC Michaelis constant for CO₂. |
| gamma_star_25, dH_gamma_star | γ* | unitless | 3.8168×10⁻⁴ / Ea=**+31.1** | 0.5 / SRubisco. Positive Ea — γ* increases with T because Rubisco specificity decreases. The von Caemmerer (2021) Setaria spreadsheet lists −31.1 (transcription error: it copied Boyd 2015's Ea for Sc/o without the required reciprocal sign flip); Woodford et al. (2025) Table 1 silently corrects to +31.1. |
| Om_25, dH_Om | Om | μbar | 210 000 / Ea=0 | Mesophyll O₂ partial pressure (ambient). |
| User-tunable scalars | ||||
| alpha_psII_fraction | α | 0..1 | 0 | Fraction of PSII activity in the bundle sheath (0 for NADP-ME; 0.5 recommended for NAD-ME / PCK species per Woodford et al. 2025). |
| x_etr_partition | x | 0..1 | 0.4 | Fraction of linear ETR partitioned to the mesophyll. |
| Vpr | Vpr | μmol/m²/s | 80 | PEP regeneration cap. |
| Rm_frac | — | unitless | 0.5 | Rm = Rm_frac · Rd. |
| fcyc | fcyc | 0..1 | 0.45 | Cyclic electron flow fraction. Updated from 0.3 (vC2021) to 0.45 (Woodford et al. 2025) reflecting NDH-dominated cyclic flow. |
| H_J | HJ | H⁺/e⁻ | 3 | Protons per electron, linear ETR (Woodford et al. 2025). |
| H_Jcyc | HJ,cyc | H⁺/e⁻ | 3.4 | Protons per electron, cyclic ETR (NDH-dominated). |
| gbs | gbs | mol/m²/s/bar | 0.003 | Bundle-sheath conductance to CO₂. |
| ao | ao | unitless | 0.047 | O₂/CO₂ solubility-diffusivity ratio. |
| absorptance | — | unitless | 0.85 | Leaf PAR absorptance. |
| f_spectral | f | unitless | 0.15 | Spectral-quality correction to absorbed PAR. |
| theta_etr | θ | unitless | 0.7 | Curvature of the J ~ I₂ non-rectangular hyperbola. |
| h_protons | — | H⁺/ATP | 4 | Protons required per ATP synthesized. |
PyHelios ships three published C4 parameter sets accessible via setC4CoefficientsFromLibrary() and getC4CoefficientsFromLibrary(). Each entry specifies every parameter the model reads — including the "complementary" kinetic constants (Kc, Ko, Kp) and scalar structural parameters that the source paper held fixed while fitting the headline rate constants. Mixing the headline values from one entry with the fixed-parameter assumptions of another will produce biased predictions, so treat each entry as atomic. Unknown species keys raise RuntimeError; key matching is case-insensitive.
| Species key | Subtype | Vcmax(25) | Vpmax(25) | Jmax(25) | Rd(25) | gm(25) | T-response | Source |
|---|---|---|---|---|---|---|---|---|
SetariaViridis_vC2021 | NADP-ME | 40 | 200 | 247.69 | 0.4 | 1.0 | Arrhenius (Jmax peaked) | von Caemmerer (2021) Table 1; kinetics Boyd et al. (2015); gm Ubierna et al. (2017); fcyc / H_Jcyc per Woodford et al. (2025). |
GenericC4_vC2000 | NADP-ME | 60 | 120 | 400 | 1.0 | ∞ (10⁴) | Q10≈2.3 → Arrhenius Ea≈61.6 | von Caemmerer (2000) Biochemical Models of Leaf Photosynthesis; plantecophys AciC4 defaults. |
Maize_Massad2007 | NADP-ME | 60 | 120 | 400 | 0.6 | ∞ (10⁴) | Peaked Arrhenius | Massad et al. (2007) Plant Cell Environ. 30:1191; plantecophys defaults at 25 °C. |
Units: μmol CO₂ / m² / s for all rate constants; mol CO₂ / m² / s / bar for gm.
Per-entry caveats:
SetariaViridis_vC2021 — the C4 struct's default constructor also carries this parameterization, except Rd: the library entry uses Rd = 0.01 · Vcmax = 0.4 μmol/m²/s per the original paper's convention, while the struct constructor sets Rd = 1.0 for historical compatibility.GenericC4_vC2000 — the original parameterization uses Q10 temperature responses. Helios approximates these with Arrhenius (Ea = R · Tref² · ln(Q₁₀)/10 with Tref = 298.15 K), matching Q10 behaviour within ~1 % over 15–40 °C. fcyc=0 recovers the linear-electron-flow ATP stoichiometry of the original vC2000 model.Maize_Massad2007 — Massad fitted the headline rate constants against Bernacchi (2001) C3-derived Kc/Ko (much smaller than the Boyd 2015 C4 values used for Setaria). **Do not substitute Setaria's Kc/Ko into this entry** — the Vcmax value is only internally consistent with the Bernacchi values. The paper assumed infinite mesophyll conductance, so the entry sets gm = 10⁴ mol/m²/s/bar as effectively infinite. The Kp temperature response is from Boyd (2015) rather than Massad's own Q10=2.1 (Massad's value overestimates Kp temperature sensitivity). The 0.6 μmol/m²/s default for Rd is a library convention (0.01 · Vcmax) — Massad themselves used Rd = 0 and reported results as insensitive to the Rd choice.Implementation notes (vs. the paper's supplementary spreadsheet):
- The Jmax temperature response uses peaked Arrhenius (same API as C3) rather than the paper's Gaussian — Jmax(25 °C) = 247.69 matches the Gaussian at 25 °C; users who need exact Gaussian behaviour at other temperatures should fit peaked-Arrhenius parameters or set Jmax as a constant.
- The Vpr cap in Eq. 19 is always applied; the paper's spreadsheet omits it, so Helios gives slightly lower A at high Cm when Vp,MM > Vpr.
For testing or validation against the von Caemmerer (2021) reference spreadsheet, setCm() lets you bypass the iterative Cm = Ci - A/gm solve and prescribe the mesophyll cytosolic CO₂ directly. The stomatal balance on Ci is also bypassed (Ci is back-computed from Ci = Cm + A/gm):
Both setC4CoefficientsFromLibrary() and setC4ModelCoefficients() accept a material_label= keyword to apply coefficients to every primitive sharing a material rather than to a UUID list. material_label and uuids are mutually exclusive:
The PhotosynthesisModel plug-in implements three types of models: the C3 biochemical model of Farquhar, von Caemmerer, and Berry (1980), the C4 biochemical model of von Caemmerer (2021), and an empirical model similar to that of Johnson (2010). Each is described separately below.
By default, the plug-in uses the C3 Farquhar model. The model can either be set explicitly, as illustrated in the code below, or the model type will be inferred based on the model coefficients that are set (see descriptions below).
The PhotosynthesisModel implements the biochemical model of Farquhar, von Caemmerer, and Berry (1980). The form used here predicts photosynthetic production as a function of photoshynthetically active radiation flux, ambient CO2 concentration, and stomatal conductance, which may itself provide responses to a number of additional environmental variables.
The implementation used here calculates the net rate of CO2 exchange as
\[A=\left(1-\frac{\Gamma^*}{C_i}\right)\mathrm{min}\left\{W_c,W_j,W_p\right\}-R_d,\]
where
\[W_c=\frac{V_{cmax}C_i}{C_i+K_{co}}\]
is the rate limited by Rubisco, with
\[K_{co}=K_c\left(1+\frac{O}{K_O}\right),\]
where \(O\) is oxygen concentration.
\[W_j=\dfrac{J C_i}{4C_i+8\Gamma^*}\]
is the rate limited by RuBP regeneration.
The light response of \(J\), the potential electron transport rate, can be described as a rectangular hyperbola with 1 shape parameter, or non-rectangular hyperbola with 2 shape parameters. The rectangular hyperbola takes the form
\[ J(Q) = \dfrac{J_{max} \alpha Q}{\alpha Q + J_{max}} \]
where \(Q\) is the photosynthetically active absorbed radiation flux ( \(\mu mol\,m^{-2}\,s^{-1}\)), \(J_{max}\) is the temperature-dependent maximum potential electron transport rate ( \(\mu mol\,m^{-2}\,s^{-1}\)), and \(\alpha\) is the intrinsic quantum efficiency of electron transport ( \(electron\,photon^{-1}\)) which defines the initial slope of the light response, determines its resulting shape in the rectangular hyperbolic form, and is thought to be relatively conserved around 0.5. The non-rectangular hyperbola takes the form
\[J(Q) = \dfrac{\alpha Q + J_{max} - \sqrt{(\alpha Q + J_{max})^2 - 4 \theta \alpha Q J_{max}}}{2 \theta}\]
where \(\theta\) is an additional parameter (unitless) that shapes the light response curve beyond its initial slope. When \(\theta\) approaches zero, the two forms become equivalent. In Photosynthesis Plugin, the rectangular form will be assumed unless \(\theta\) is specified by the user.
\[W_p=\dfrac{3\,TPU\,C_i}{C_i-\Gamma^*}\]
is the rate limited by triose phosphate utilization. Note that if the TPU parameter is not set by the user, this state is ignored.
The intercellular CO2 concentration \(C_i\) is determined from the relation
\(A=0.75g_M\left(C_{a}-C_i\right)\)
which is solved numerically using the Secant method, since \(A\) is a complex nonlinear function of \(C_i\) which prevents an analytical solution for \(C_i\). The 0.75 factor comes from the fact that diffusion of CO2 in air is slower than that of water vapor (see Eq. 7.33 of Campbell and Norman).
\(g_M\) is the conductance to moisture transfer between the leaf interior and just outside of the leaf boundary-layer, and is calculated as
\(g_M = 1.08g_Hg_S\left[\dfrac{\zeta}{1.08g_H+g_S\zeta}+\dfrac{(1-\zeta)}{1.08g_H+g_S(1-\zeta)}\right]\),
where \(g_H\) is the boundary-layer conductance to heat, and \(1.08g_H\) gives the boundary-layer conductance to moisture considering the differences in diffusivity between water vapor and heat. \(n_s\) is the number of primitive faces, which is determined by the value of "twosided_flag" retrieved from the primitive's assigned material when available (see Context::getMaterialTwosidedFlag()), with fallback to primitive data if no user-assigned material exists (twosided_flag=0 is single-sided and \(n_s=1\), twosided_flag=1 is two-sided and \(n_s=2\)).
\(\zeta=\dfrac{D_{upper}}{D_{lower}+D_{upper}}\)
is the stomatal sidedness, which is the ratio of the stomatal density of the upper leaf surface to the sum of the upper and lower leaf surface densities, which is set by the primitive data value "stomatal_sidedness". For leaves, \(\zeta=0\) corresponds to hypostomatous leaves (stomata on one side), and \(\zeta=0.5\) to amphistomatous leaves (stomata equally on both sides). It is important to note that if \(n_s=1\), then the value of \(\zeta\) will be overridden and set to 0.
By default the implementation assumes the chloroplast CO2 concentration equals the intercellular value, \(C_c \equiv C_i\) (infinite mesophyll conductance). A finite mesophyll conductance \(g_m\) (mol CO2 m-2 s-1 bar-1) can be supplied to model diffusion from the intercellular airspace to the sites of carboxylation:
\[ C_c = C_i - \dfrac{A}{g_m}. \]
Substituting \(C_c\) into the Rubisco-limited and electron-transport-limited forms of \(A\) gives two quadratic equations in the net assimilation, one per branch, that are solved analytically following Ethier & Livingston (2004) and Sharkey et al. (2007). The physically meaningful (smaller) root is selected and combined with \(A_p = 3 \cdot TPU - R_d\) via the same smooth-min used elsewhere.
gm has the same temperature-response options as every other rate parameter: constant, Arrhenius, peaked-Arrhenius (with default dHd), and peaked-Arrhenius with explicit dHd. The Python wrapper collapses the four C++ overloads into a single method whose unused arguments default to the -1 sentinel:
The default value gm = +∞ reproduces the legacy Farquhar Cc = Ci behaviour bit-for-bit, so existing parameter libraries that do not set gm are unaffected.
Two different temperature response functions are commonly used in photosynthetic modeling and supported in the Photosynthesis Plugin. One is an Arrhenius equation, which is exponentially increasing with no decline in the region of use. The other is a modified Arrhenius equation with a peak or temperature optimum, beyond which there is a decline in the value of the function, representing a denaturing of an enzyme and subsequent reduction of its activity.
\[ \begin{aligned} k &= k_{25} \cdot \exp \left[\frac{\Delta H_a}{R} \left(\frac{1}{298}-\frac{1}{T_{leaf}} \right) \right] \frac{f(298)}{f(T_{leaf})}, \\ f(T_{leaf}) &= 1+\exp \left[\frac{\Delta H_d}{R} \left(\frac{1}{T_{opt}} - \frac{1}{T_{leaf}} \right) - \ln \left(\frac{\Delta H_d}{\Delta H_a}-1 \right) \right] \end{aligned} \]
In this form, the model is conveniently parameterized by the commonly used standard reference rate at 25 \(^\circ\)C, \(k_{25}\), as well as the energy of activation, \(A = \Delta H_a = dH_a\) of the Arrhenius equation, but also by the observable temperature optimum, \(T_{opt}\) and one additional fitted parameter, the energy of deactivation, \(D = \Delta H _d = dH_d\), related to the rate of decline from the optimum.
As \(T_{opt}\) \(\to \infty\), then the peaked form approaches the standard, unpeaked Arrhenius equation, allowing for mathematical backwards compatibility for parameters obtained from fitting to the standard unpeaked form.
In the Photosynthesis Plugin, the Arrhenius form will be assumed as it requires fewer parameters, unless the additional parameters \(dH_d\) and \(T_{opt}\) are specified by the user.
| Parameter | Description | Units |
|---|---|---|
| \(k_{25}\) | reference rate at 25 \(^\circ\)C | \(\mu mol\,m^{-2}\,s^{-1}\) |
| \(dH_a\) | activation energy (rate of increase) | \(kJ\,mol^{-1}\) |
| \(dH_d\) | deactivation energy (rate of decrease) | \(kJ\,mol^{-1}\) |
| \(T_{opt}\) | optimum temperature in Kelvin | \(K\) |
| \(T_{leaf}\) | leaf surface temperature in Kelvin | \(K\) |
| \(R\) | ideal gas constant, 0.008314 | \(kJ\,mol^{-1}\,K^{-1}\) |
Additional temperature parameters that are not typically fit to use the standard Arrhenius form with the parameters obtained by Bernacchi et al. (2001), and are given by the following equations
\[ \begin{aligned} \Gamma^* &= 42.60 \cdot \exp \left[ \frac{37.83}{R} \left(\frac{1}{298} - \frac{1}{T_{leaf}} \right) \right] \\ K_c &= 400.3 \cdot \exp \left[ \frac{79.43}{R} \left(\frac{1}{298} - \frac{1}{T_{leaf}} \right) \right] \\ K_o &= 275.1 \cdot \exp \left[ \frac{36.38}{R} \left(\frac{1}{298} - \frac{1}{T_{leaf}} \right) \right] \\ R_d &= R_{d,25} \cdot \exp \left[ \frac{46.39}{R} \left(\frac{1}{298} - \frac{1}{T_{leaf}} \right) \right] \end{aligned} \]
| Variable | Units | Description |
|---|---|---|
| \(Q\) | \(\mu\)mol/m2-sec. | Photosynthetic radiation energy flux. |
| \(T_s\) | Kelvin | Surface temperature. |
| \(C_{a}\) | \(\mu\)mol CO2/mol air | Ambient CO2 concentration outside of boundary-layer. |
| \(g_M\) | mol air/m2-s | Conductance to moisture transfer between inside the leaf and leaf surface (i.e., stomatal conductance). |
| \(g_H\) | mol air/m2-s | Conductance to heat transfer between the leaf surface and outside the boundary-layer (i.e., boundary-layer conductance). |
The table below gives example model parameters obtained for several different species. These parameters were fit from leaf-level gas exchange data using the PhoTorch Python package. Note that the parameter sets are based on different temperature response functions depending on the data that was available.
| Species | \(V_{cmax25}\) | \(J_{max25}\) | \(TPU_{25}\) | \(R_{d25}\) | \(\alpha\) | \(\theta\) | \(\Delta H_{a,Vcmax}\) | \(T _{opt,Vcmax}\) | \(\Delta H_{d,Vcmax}\) | \(\Delta H_{a,Jmax}\) | \(T _{opt,Jmax}\) | \(\Delta H_{d,Jmax}\) | \(\Delta H_{a,TPU}\) | \(T _{opt,TPU}\) | \(\Delta H_{d,TPU}\) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Almond | 105.9 | 166.34 | – | 1.49 | 0.336 | 0 | 65.33 | – | – | 46.36 | – | – | – | – | – |
| California Bay | 97.5 | 193 | 3.3 | 0.1 | 0.037 | 0 | 49.1 | 308.6 | 505.8 | 34 | 308.5 | 456.7 | 0.1 | 309.4 | 477.5 |
| Elderberry | 37.7 | 149.7 | 7.3 | 1.3 | 0.202 | 0.472 | 66 | 319.4 | 496 | 24.5 | 314.8 | 492.9 | 33.6 | 314.5 | 497.5 |
| Grape | 74.5 | 180.2 | 7.7 | 1.3 | 0.304 | 0 | 76.1 | 318.8 | 499.8 | 23 | 313.8 | 502.3 | 24 | 314.6 | 496.4 |
| Maple | 96.4 | 168 | 2.7 | 0.1 | 0.077 | 0 | 48.9 | 307.1 | 505 | 8.5 | 304.7 | 476.7 | 32.1 | 308.3 | 471.6 |
| Olive | 75.9 | 170.4 | 8.3 | 1.9 | 0.398 | 0 | 55.4 | 315.2 | 497 | 32.2 | 312.5 | 493.4 | 37.2 | 311.7 | 498.9 |
| Pistachio (female) | 138.99 | 221.76 | – | 2.85 | 0.366 | 0 | 65.33 | – | – | 43.80 | – | – | – | – | – |
| Pistachio (male) | 154.17 | 243.20 | – | 2.05 | 0.335 | 0 | 65.33 | – | – | 50.89 | – | – | – | – | – |
| Toyon | 52.8 | 142.4 | 6.6 | 0.8 | 0.29 | 0.532 | 42.1 | 315.1 | 483 | 9 | 313 | 486.2 | 14 | 314.8 | 493.8 |
| Walnut | 121.85 | 197.25 | – | 1.96 | 0.404 | 0 | 65.33 | – | – | 48.35 | – | – | – | – | – |
| Redbud | 68.5 | 132.4 | 6.6 | 0.8 | 0.41 | 0.04 | 66.6 | 315.1 | 496 | 41.2 | 313.1 | 474 | 34.3 | 312.8 | 463.2 |
| Apple | 101.08 | 167.03 | – | 3.00 | 0.432 | 0 | 65.33 | – | – | 47.62 | – | – | – | – | – |
| Cherry | 75.65 | 129.06 | – | 2.12 | 0.404 | 0 | 65.33 | – | – | 48.49 | – | – | – | – | – |
| Pear | 107.69 | 176.71 | – | 1.51 | 0.274 | 0 | 65.33 | – | – | 46.04 | – | – | – | – | – |
| Prune | 75.88 | 129.41 | – | 1.65 | 0.402 | 0 | 65.33 | – | – | 48.58 | – | – | – | – | – |
The model coefficients can be set manually or by using the library of coefficients provided in the table above.
To load the FvCB parameters from the library, call the function setFarquharCoefficientsFromLibrary() with the species name as an argument. This will automatically set the parameters for all primitives. Alternatively, a list of UUIDs can be passed to this function to set the parameters for a subset of primitives.
To set the parameters manually, first create an instance of the FarquharModelCoefficients class, and then set the parameters using the setter methods. Finally, call the setFarquharModelCoefficients() method with the coefficients object as an argument.
Each parameter has a setter function, which is the means by which the underlying response function to be used is specified.
Parameter Temperature Response
Light Response
Note that the parameter sets in the library vary in terms of which response functions are used based on the data available for parameter fitting.
The native Helios C++ library uses a material-based approach for setting photosynthesis model coefficients:
C++ Approach (Native Helios):
Advantages of material-based approach (C++):
PyHelios Implementation Note:
PyHelios currently uses a UUID-based approach rather than the material-based system. Pass a list of primitive UUIDs to set coefficients for specific primitives:
The UUID-based approach provides similar functionality, allowing you to group primitives logically and set parameters per group.
The PhotosynthesisModel also implements an empirical photosynthesis model using EmpiricalModelCoefficients. The net photosynthetic rate is described by the equation:
\(A = A_{sat} f_L f_T f_C - R_d\)
\(A_{sat}\,({\mu}mol/m^2-s)\) is the photosynthesis assimilation rate at saturating irradiance and reference temperature ( \(T_{ref}\)) and intercellular CO2 concentration ( \(C_{i,ref}\)).
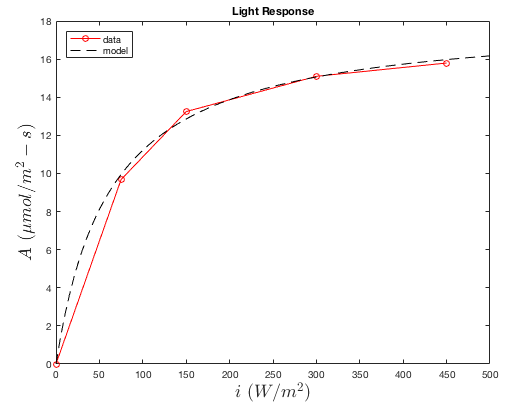
The response of photosynthesis to light is given by a simple exponential function, which is defined by only one parameter:
\(f_L(i) = \dfrac{i}{\theta+i}\),
where \(\theta\) is the light response curvature.
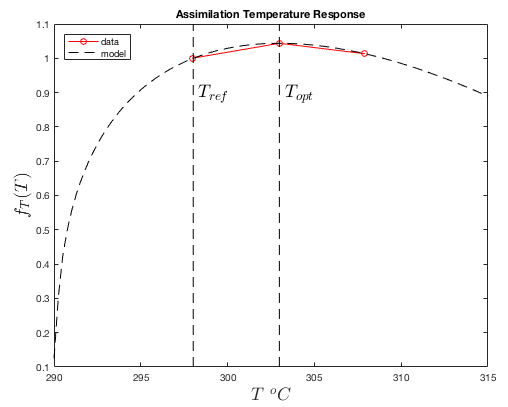
It is assumed that the maximum CO2 assimilation rate \(A_{max}\) decreases exponentially about some optimum temperature \(T_{opt}\). The temperature response function is given by:
\(f_T(T_s) = \left(\dfrac{T_s-T_{min}}{T_{ref}-T_{min}}\right)^q\left(\dfrac{(1+q)T_{opt}-T_{min}-qT_s}{(1+q)T_{opt}-T_{min}-qT_{ref}}\right)\),
where \(T_{min}\) is the minimum temperature at which assimilation occurs, \(T_{opt}\) is the temperature at which the maximum assimilation rate occurs, \(T_{ref}\) is the reference temperature chosen to define \(A_{ref}\), and \(q\) is a shape parameter.
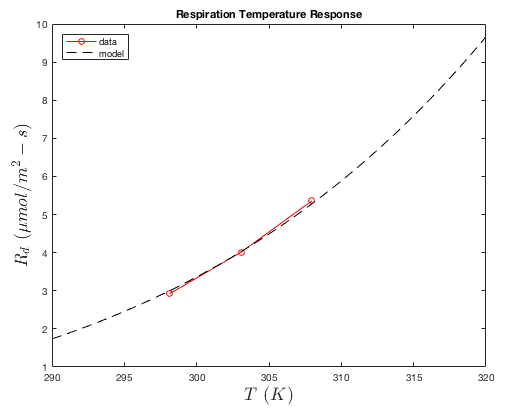
The "dark" respiration rate \(R_d\) is assumed to increase exponentially with temperature following the Arrhenius equation (and assumed not to vary with ambient CO2 concentration). Thus, the dark respiration rate is calculated simply as
\(R_d = R\sqrt{T_s-273}\;\mathrm{exp}\left(-E_R/T_s\right)\),
where \(R\) and \(E_R\) are parameters, and \(T_s\) is the surface temperature in Kelvin (the \(-273\) inside the square root converts to degrees Celsius).
We assume that the maximum assimilation rate varies linearly with intercellular CO2 concentration over the range of expected concentrations, and is zero at zero CO2. Thus, the response function is simply
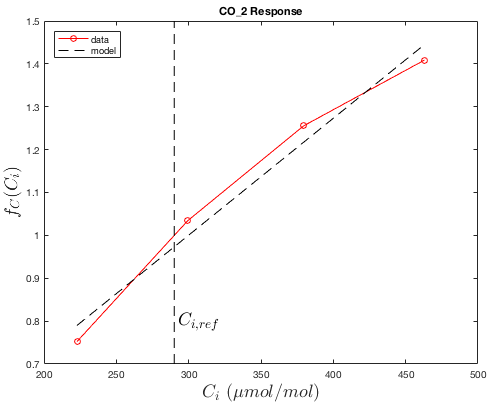
\(f_C(C_i) = k_C\dfrac{C_i}{C_{i,ref}}\),
where \(C_i\) is intercellular CO2 concentration ( \(\mu\)mol CO2/mol air).
The intercellular CO2 concentration is estimated as a function of the boundary-layer conductance, stomatal conductance, and ambient CO2 concentration outside of the primitive boundary-layer. The rate of transport of CO2 to the leaf (i.e., assimilation rate) is given by
\(A = 0.75g_M\left(C_{amb}-C_i\right)\),
where \(g_M\) is the conductance to moisture from the sub-stomatal cells to outside of the boundary-layer. The 0.75 factor comes from the fact that diffusion of CO2 in air is slower than that of water vapor (see Eq. 7.33 of Campbell and Norman).
Since \(A\) is dependent on \(C_i\) and vice-versa, an iterative solution is required for \(A\).
| Variable | Units | Description |
|---|---|---|
| \(i\) | W/m2 | Photosynthetic radiation energy flux. |
| \(T_s\) | Kelvin | Surface temperature. |
| \(C_{amb}\) | \(\mu\)mol CO2/mol air | Ambient CO2 concentration outside of boundary-layer. |
| \(g_{bl}\) | mol air/m2-s | Boundary-layer conductance. |
| \(g_s\) | mol air/m2-s | Stomatal conductance. |
| Parameter | Units | Description |
|---|---|---|
| \(A_{sat}\) | mol CO2/m2-sec | Assimilation rate at saturating irradiance and reference temperature and intercellular CO2 concentration. |
| \(\theta\) | W/m2 | Shape parameter for response to light. |
| \(T_{min}\) | Kelvin | Minimum temperature at which assimilation occurs. |
| \(T_{opt}\) | Kelvin | Temperature at which maximum assimilation rate occurs. |
| \(q\) | unitless | Temperature response shape function. |
| \(R\) | \({\mu}\)mol K1/2/m2-s | Pre-exponential factor for respiration temperature response. |
| \(E_R\) | 1/Kelvin | Respiration temperature response rate. |
| \(k_C\) | unitless | CO2 response rate. |
The response of the assimilation rate to light is obtained from gas exchange measurements at reference temperature ( \(T_{ref}\)) and CO2 ( \(C_{i,ref}\)) in which the irradiance is varied across some range. However, one important detail is that the dark respiration rate should be removed such that \(A=0\) in the dark (see plot below). This can be done by measuring the net CO2 flux starting in the dark, then subtracting the dark flux from the total flux for each subsequent light level.
The response of the assimilation rate to temperature is obtained using gas exchange measurements at saturating light levels and the reference CO2 concentration. The temperature is varied across some range, and the assimilation rate is measured. It is assumed that the optimum temperature \(T_{opt}\) is the temperature corresponding to the maximum measured assimilation rate. The model is fit to the data to determine \(T_{min}\) and \(q\).
The response of the dark respiration to temperature is obtained using gas exchange measurements in the dark. The leaf is first acclimated to the dark chamber, then leaf temperature is varied across some range. The model is then fit to the data to determine parameters.
The response of the assimilation rate is obtained using gas exchange measurements at saturating light levels and the reference temperature \(T_{ref}\), but with varying external CO2 concentration (which produces varying intercellular CO2).